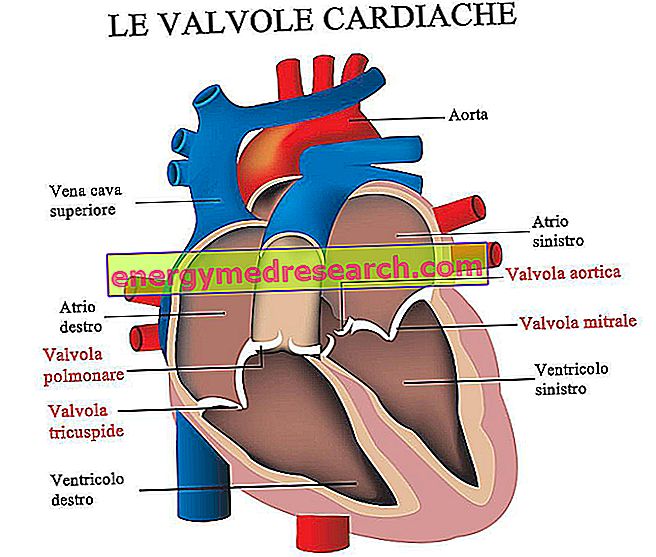
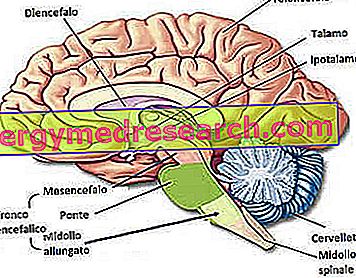
Dr Francesco Grazzina
Skeletilihase poolt kokkutõmbumise ajal tekkinud jõud on tingitud keerulisest sündmuste seeriast, mille kompromiss võib mis tahes tasemel kaasa aidata neuromuskulaarse väsimuse tekkele.
Selleks, et lihaskiud saaks kokku leppida, peab depolariseerimise impulss jõudma seljaajumootori neuronist.

Eksperimentaalselt on väsimus jagatud "keskseks" ja "perifeerseks".
Keskne väsimus ja perifeerne väsimus
Väsimus on määratletud kui "keskne", kui see on tingitud mehhanismidest, mis pärinevad kesknärvisüsteemi tasemest, või pigem nendest struktuuridest, mille ülesanded ulatuvad liikumise kontseptsioonist kuni närviimpulsside juhtumiseni seljaajumootori neuronisse. Seda määratletakse kui "perifeerset väsimust", kui nähtused, mis seda määravad, esinevad seljaajureaktoris, mootori naastul või skeleti lihasrakus.
Seetõttu on keskne väsimus väljendus neuronite liikumise vähenemisest skeletilihastele. Kuid kesknärvisüsteemi aktiveerimise taset saab suurendada, kui subjekt on asjakohaselt stimuleeritud verbaalse julgustamise või erinevat tüüpi tagasisidega. Seega mängiks keskne süsteem väsimuse tekkimisel otsustavat rolli.
Mis puudutab sporti, siis tuleb öelda, et kesksed tegurid, nagu psühholoogiline motivatsioon, emotsionaalne enesekontroll ja füüsilise ebamugavuse tolerantsus, mängivad keerulises lihasaktiivsuses, mis on sportliku žesti aluseks.
Praegused uuringud näitavad, et väsimuse alguskoht on lihases ja seetõttu soodustavad nad väsimuse perifeerset paiknemist. Anatoomilised struktuurid, mis võivad kaasa aidata lokaliseeritud lihaste väsimuse kujunemisele, on seljaajuraku neuron, neuromuskulaarne ühendus, sarkolemma ja lihaskiudude T-süsteem.
Teine tegur, millest sõltub väsimus, on tasakaalustamatus ATP kasutamise kiiruse ja sünteesikiiruse vahel. Tegelikult ei ole oluline selle vaba energia doonori koguhulk, vaid pigem Pi, mis vabaneb ATP hüdrolüüsi teel. Tõepoolest tundub, et selle suurenemine vähendab astiin-müosiini sildade moodustumist, mis takistab kontraktiilset mehhanismi.
Lihas-glükogeeni kättesaadavus muutub oluliseks harjutuste puhul, mis nõuavad hapniku tarbimist 65% kuni 85% ulatuses maksimaalsest hapnikutarbimisest, mida toetavad peamiselt II tüüpi väsimuskindlad kiud.
Kõrgema intensiivsusega harjutuste puhul esindavad energiaallikaid peamiselt ringlev glükoos. Maksimaalse intensiivsuse harjutused katkestavad piimhappe suurenemine enne, kui lihaste glükogeeni tase saavutab jõudlust piiravad väärtused.