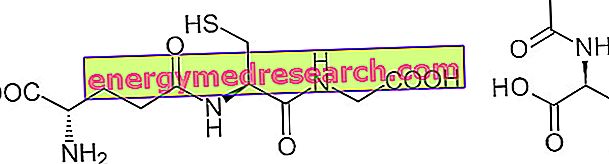
Et rääkida kahekümnest aminohappest, mis moodustavad valku ja modifitseeritud struktuure, tuleb kirjeldada vähemalt 12 erialast metaboolset rada.
Aga miks rakud kasutavad aminohapete kataboliseerimiseks nii palju metaboolseid teid, mis vajavad energiat (näiteks ensüümide katalüütiliste saitide regenereerimiseks)? Peaaegu kõiki aminohappeid on võimalik saada spetsiaalsete marsruutide kaudu metaboliitide abil, mida väikese osa kasutatakse energia tootmiseks (näiteks glükoneogeneesi ja ketoonkehade teed), kuid mis eelkõige põhjustavad komplekssete molekulide moodustumist, \ t suure hulga süsinikuaatomitega (näiteks fenüülalaniinist ja türosiinist, toodetakse sellel eesmärgil spetsialiseerunud neerupealistes hormoonid); kui ühest küljest oleks lihtne toota energiat aminohapetest, teisest küljest oleks keeruline ehitada keerulisi molekule, alustades väikestest molekulidest: aminohapete katabolism võimaldab meil kasutada oma luustikku suuremate liikide saamiseks.
Kaks või kolm untsi aminohappeid lagunevad terve inimese poolt iga päev: 60-100 g neist pärineb dieedist toodetud valkudest, kuid üle 2 untsi saadakse keha lahutamatuks osaks olevate valkude normaalsest käibest (aminohapped) nendest valkudest, mida redoksprotsessid kahjustavad, asendatakse teised ja kataboliseeruvad).
Aminohapped annavad energiasisalduse ATP järgi: pärast a-aminorühma eemaldamist võib aminohapete järelejäänud süsiniku skelett pärast asjakohaseid transformatsioone siseneda krebitsüklisse. Lisaks sellele, kui toitainete kogusest puudub ja glükoosi kogus väheneb, aktiveeritakse glükoneogenees: glükoneogeneetilisi aminohappeid nimetatakse neile, mis pärast asjakohaseid modifikatsioone saab glükoneogeneesi sisse viia; glükoneogeneetilised aminohapped on need, mida saab muundada püruvaadiks või fumaraadiks (fumaraati saab muundada haigeks, mis väljub mitokondritest ja tsütoplasmas transformeeritakse oksaloatsetaadiks, millest saadakse fosfolool). Selle asemel väidetakse, et need, mida saab muuta atsetüülkoensüümiks A ja äädika-atsetaadiks, on ketogeensed aminohapped.
Kirjeldatud on väga oluline aspekt, sest aminohapped võivad kohe tühja kõhuga suhkru puudumise kõrvaldada; kui paastumine püsib, sekkub lipiidide ainevahetus kahe päeva pärast (kuna valgu struktuure ei saa palju rünnata), siis selles etapis, kuna glükoneogenees on väga piiratud, muudetakse rasvhapped atsetüülkoensüüm A ja ketoonkehadeks . Edasisest paastumisest kohandub aju ka ketoonkehade kasutamisel.
A-aminorühma ülekandmine aminohapetest toimub läbi transamiinimisreaktsiooni; nimetatud reaktsiooni katalüüsivaid ensüüme nimetatakse transaminaasideks (või amino transferaasideks). Need ensüümid kasutavad ensümaatilist kofaktorit, mida nimetatakse püridoksaalfosfaadiks, mis sekkub selle aldehüüdi rühmaga. Püridoksaalfosfaat on püridoksiini fosforüülimise saadus, mis on peamiselt köögiviljades sisalduv vitamiin (B6).
Transaminaasidel on järgmised omadused:
A-ketoglutaraadi-glutamaadi paari kõrge spetsiifilisus;
Nad võtavad oma nime teiselt paarilt.
Transaminaasi ensüümid hõlmavad alati a-ketoglutaraadi-glutamaadi paari ja neid eristab teine kaasatud paar.
näited:
Aspartaat-transaminaas või GOT (glutamaadi-oksaalatsetaadi transaminaas): ensüüm kannab a-aminorühma aspartaadist a-ketoglutaraadile, saades oksalatsetaadi ja glutamaadi.
Alaniini transaminaas, st GTP (glutamaat-püruvaadi transaminaas): ensüüm kannab a-aminorühma alaniinist a-ketoglutaraadile, saades püruvaadi ja glutamaadi.
Erinevad transaminaasid kasutavad aminohapete aminorühma aktseptorina a-ketoglüraati ja muundavad selle glutamaadiks; samal ajal kasutatakse moodustunud aminohappeid ketoonkehade teel.
Selline reaktsioon võib toimuda mõlemas suunas, kuna need lagunevad ja moodustuvad sama energiasisaldusega sidemed.
Transaminaasid on nii tsütoplasmas kui ka mitokondrites (need on enamasti tsütoplasmas aktiivsed) ja erinevad isoelektrilisest punktist.
Transaminaasid on samuti võimelised dekarboksüülima aminohappeid.
Glutamaadi muundamiseks α-ketoglutaraadiks peab olema võimalus: see toimub deaminatsiooni teel.
Glutamaadi dehüdrogenaas on ensüüm, mis on võimeline transformeerima glutamaati a-ketoglutaraadiks ja seega muundama aminohapete aminohapped, mis on leitud glutamaadi vormis, ammoniaagiks. Mis juhtub, on oksüdoreduktiivne protsess, mis läbib a-amino-glutaraadi vaheprodukti: vabaneb ammoniaak ja a-ketoglutaraat ning pöörduvad ringlusse.
Seega läbib aminohapete aminorühmade kõrvaldamine läbi transaminaaside (erinevad sõltuvalt substraadist) ja glutamaadi dehüdrogenaasist, mis määrab ammoniaagi moodustumise.
Glutamaadi dehüdrogenaasi on kahte tüüpi: tsütoplasmaatiline ja mitokondriaalne; kofaktor, mis on ka selle ensüümi kaasosa, on NAD (P) +: glutamaadi dehüdrogenaas kasutab redutseeriva võimsuse aktseptorina NAD + või NADP +. Tsütoplasma vorm aga eelistab, kuigi mitte ainult, NADP +, samas kui mitokondriaalne vorm eelistab NAD +. Mitokondriaalse vormi eesmärk on kõrvaldada aminorühmad: see põhjustab ammoniaagi (mis on spetsiifilise mitokondriumi ensüümi substraat) ja NADH (mis saadetakse hingamisteedesse) moodustumist. Tsütoplasma vorm toimib vastupidises suunas, see tähendab, et ta kasutab ammoniaaki ja a-ketoglutaraati, et saada glutamaati (millel on biosünteetiline sihtkoht): see reaktsioon on redutseeriv biosüntees ja kasutatud kofaktor on NADPH.
Glutamaadi dehüdrogenaas toimib siis, kui aminohapete nagu ammoniaak (uriini kaudu) aminorühmad tuleb kõrvaldada või kui energiatootmiseks on vaja aminohapete skelette: sellel ensüümil on seega süsteemid, mis osutavad negatiivseks modulaatoriks head energia kättesaadavust (ATP). GTP ja NAD (P) H) ja positiivsete modulaatoritena süsteemid, mis viitavad energiavajadusele (AMP, ADP, SKP, NAD (P) +, aminohapped ja kilpnäärme hormoonid).
Aminohapped (peamiselt leutsiin) on glutamaadi dehüdrogenaasi positiivsed modulaatorid: kui tsütoplasmas esineb aminohappeid, võib neid kasutada valgusünteesiks või need tuleb kõrvaldada, sest neid ei saa koguda (sel põhjusel on aminohapped positiivsed modulaatorid). .
Ammoniaagi kõrvaldamine: uurea tsükkel
Kala kõrvaldab ammoniaagi, viies selle vee kaudu läbi küünte; linnud muudavad selle kusihappeks (mis on kondensatsiooniprodukt) ja kõrvaldavad selle väljaheitega. Vaatame, mis juhtub inimestega: oleme öelnud, et glutamaadi dehüdrogenaas muundab glutamaadi α-ketoglutaraadiks ja ammoniaagiks, kuid me ei ole öelnud, et see juhtub ainult maksa mitokondrites.
Ammoniaagi kõrvaldamise põhiline roll karbamiiditsükli kaudu on kaetud mitokondriaalsete transaminaasidega.

süsinikdioksiidi, bikarbonaadi ioonina (HCO3-), aktiveerib biotiini kofaktor, mis moodustab karboksübiotiini, mis reageerib ammoniaagiga, saades karbaamhappe; järgnev reaktsioon kasutab ATP-d fosfaadi ülekandmiseks karbaamhappesse, moodustades karbamüülfosfaadi ja ADP-d (ATP konversioon ADP-ks on karboksibiotiini saamise mootor). Seda faasi katalüüsib karbamüülfosfaadi süntetaas ja see toimub mitokondrites. Karbamüülfosfaat ja ornitiin on substraadid ornitiini trans-karbamasülaasi ensüümile, mis muundab need tsitrulliiniks; see reaktsioon esineb mitokondrites (hepatotsüütides). Toodetud tsitrulliin väljub mitokondritest ja tsütoplasmas läheb arginiinsuktsinaadi süntaasi toimele: tsitruliini süsiniku skeemi ja aspartaadi vahel on liitumine nukleofiilse rünnaku ja sellele järgneva vee kõrvaldamise vahel. Ensüüm arginiinsuktsinaadi süntaas nõuab ATP-molekuli, seega on olemas energiline sidestus: ATP hüdrolüüs AMP-ks ja pürofosfaadiks (viimane muundatakse seejärel kaheks ortofosfaatmolekuliks), eraldades molekuli. veest substraadist, mitte vee toimest keskkonda.
Järgmine ensüüm on arginiini suktsinaas : see ensüüm on võimeline jagama arginiini suktsinaati arginiiniks ja fumaraadiks tsütoplasmas.
Uureatsükkel viiakse lõpule ensüümi arginaasiga : saadakse karbamiid ja ornitiin; uurea kõrvaldatakse neerude poolt (uriin), samas kui ornitiin naaseb mitokondritesse ja jätkab tsüklit.
Karbamiiditsükkel toimub arginiini kaudu kaudselt: arginiini akumulatsioon näitab, et on vaja kiirendada uurea tsüklit; arginiini modulatsioon on kaudne, sest arginiin moduleerib positiivselt atsetüülglutamaadi süntetaasi ensüümi. Viimane on võimeline kandma atsetüülrühma glutamaadi lämmastikule: moodustub N-atsetüülglutamaat, mis on ensüümi karbamüülfosfosüntetaasi otsene modulaator.
Arginiin koguneb karbamiidtsükli metaboliidina, kui karbamüülfosfaadi tootmine ei ole ornitiini kõrvaldamiseks piisav.
Karbamiid toodetakse ainult maksas, kuid on ka teisi alasid, kus toimub algne reaktsioon.
Aju ja lihased kasutavad aminorühmade kõrvaldamiseks spetsiaalseid strateegiaid. Aju kasutab väga tõhusat meetodit, milles kasutatakse ensüümi glutamiini süntetaasi ja ensüümi glutamaasi : esimene on neuronites, viimane aga maksas. See mehhanism on väga tõhus kahel põhjusel:
Kaks aminorühma transporditakse ajust maksasse ainult ühe vehiikliga;
Glutamiin on palju vähem mürgine kui glutamaat (glutamaat kannab ka neuronaalset ülekannet ja ei tohi ületada füsioloogilist kontsentratsiooni).
Kala puhul toob sarnane mehhanism aminohapete aminorühma küünaldesse.
Lihasest (skeleti ja südamest) jõuavad aminorühmad maksas glükoosi-alaniini tsükli kaudu; kaasatud ensüüm on glutamiin-püruvaadi transaminaas: see võimaldab aminorühmade (mis on glutamaadi kujul) ülevõtmist, muundades püruvaadi alaniiniks ja samaaegselt glutamaadiks a-ketoglutaraadiks lihastes ja katalüüsides pöördprotsessi maksas.
Erinevate ülesannete või positsioonidega transaminaasidel on ka struktuursed erinevused ja neid saab määrata elektroforeesiga (neil on erinevad isoelektrilised punktid).
Transaminaaside esinemine veres võib olla maksa- või kardiopaatilise kahjustuse sümptom (st maksa- või südamerakkude kahjustus); transaminaaside kontsentratsioonid on nii maksas kui ka südames väga kõrged: elektroforeesi abil saab kindlaks teha, kas kahjustus esines maksa- või südamerakkudes.