Hemoglobiini (Hb) põhjalikuks rääkimiseks on kasulik käsitleda kõigepealt müoglobiini (Mb), mis on väga sarnane hemoglobiiniga, kuid on palju lihtsam. Hemoglobiini ja müoglobiini vahel on ranged sugulussuhted: mõlemad on konjugeeritud valgud ja nende proteesirühm (mitte-valgu osa) on hemorühm .
Müoglobiin on globulaarne valk, mis koosneb ühest kuni viiekümne aminohappe ahelast (sõltuvalt organismist) ja selle molekulmass on umbes 18 Kd.
Nagu mainitud, on see varustatud heme rühmaga, mis sisestatakse valgu hüdrofoobse (või lipofiilsesse) ossa, mis koosneb kiududest, mis on omistatavad kiuliste valkude a-heeliksi struktuuridele.
Müoglobiin koosneb peamiselt α-heelide segmentidest, mis esinevad kaheksas ja mis koosneb peaaegu eranditult mittepolaarsetest jääkidest (leutsiin, valiin, metioniin ja fenüülalaniin), samas kui polaarsed jäägid praktiliselt puuduvad (asparagiinhape, glutamiinhape, lüsiin ja arginiin); ainsad polaarsed jäägid on kaks histidiini, mis mängivad olulist rolli hapniku rünnakus hemirühmale.
Heme rühm on kromofoorirühm (imendub nähtavasse) ja on müoglobiini funktsionaalne rühm.
Vt ka: glükeeritud hemoglobiin - hemoglobiin uriinis
Väike keemia
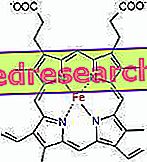
Seos protoporfüriini ja raua vahel on nn koordineerivate ühendite tüüpiline side, mis on keemilised ühendid, milles tsentraalne aatom (või ioon) moodustab sidemeid teiste keemiliste liikidega suuremal arvul kui selle oksüdatsiooninumber (elektrilaeng). Heme puhul on need võlakirjad pöörduvad ja nõrgad.
Raua koordinaatide arv (koordinaatide arv) on kuus: rauda ümber võib olla kuus molekuli, mis jagavad sidumiselektoneid.
Koordineeriva ühendi moodustamiseks kulub korrektse orientatsiooniga kaks orbitaali: üks on võimeline ostma elektrone ja teine suudab neid annetada
Heemis moodustab raud nelja lämmastikuaatomiga nelja tasapinnalise sideme, mis asuvad proto-porfüriinitsükli keskel, ja viies side proksimaalse histidiini lämmastikuga; raual on kuues vaba koordineerimislüli ja võib seonduda hapnikuga.
Kui raud on vaba iooni kujul, on kõigil d- tüüpi orbitaalidel sama energia; müoglobiini korral seondub raudioon protoporfüriiniga ja histidiiniga: need liigid häirivad magnetiliselt raud raudteed; häirete ulatus on erinevates orbitaalides erinev nende ruumilise orientatsiooni ja häirivate liikide järgi. Kuna orbitaalide koguenergia peab naasma konstantse tasemeni, põhjustab häire energiapõhise eraldumise erinevate orbitaalide vahel: mõnede orbitaalide poolt omandatud energia on võrdne teiste kaotatud energiaga.
Kui orbitaalide vaheline eraldumine ei ole väga suur, on eelistatud kõrge spin-elektrooniline paigutus: siduvad elektronid püüavad korraldada paralleelsed keerdud mitmetes võimalikes alamtasemetes (maksimaalne mitmekesisus); teisest küljest, kui häirimine on väga tugev ja orbitaalide vahel on suur eraldatus, võib olla mugavam siduda sidumiselektronid madalamate energia orbitaalidega (madal spin).
Kui raud seondub hapnikuga, võtab molekul väikese tsentrifuugimise, samas kui raual on kuues vaba koordineerimislüli, on molekulil suur spin.
Tänu sellele erinevusele spinil, suudame müoglobiini spektraalanalüüsi abil aru saada, kas on seotud hapnik (MbO2) või mitte (Mb).
Müoglobiin on lihastele iseloomulik valk (kuid mitte ainult lihastes).
Müoglobiin ekstraheeritakse sperma vaalast, milles seda esineb suurtes kogustes ja seejärel puhastatakse.
Vaalaliste hingamine sarnaneb inimestega: kopsud peavad imenduma õhku hingamisteede kaudu; sperma vaal peab sisaldama nii palju hapnikku kui võimalik lihastes, mis on võimelised hapnikku kogunema, sidudes need müoglobiiniga; hapnik vabastatakse seejärel aeglaselt, kui vaalaja on kastetud, sest selle ainevahetus nõuab hapnikku: mida suurem on hapniku kogus, mida sperma vaal suudab absorbeerida ja mida rohkem hapnikku sukeldumise ajal on saadaval.

Liha rohkem või vähem punaseks muutmiseks on hemoproteiinide sisaldus (see on liha, mis muudab liha punaseks).
Hemoglobiinil on müoglobiiniga palju struktuurset analoogiat ja ta on võimeline molekulaarset hapnikku tagasi pöörduma; kuid kuigi müoglobiin piirdub üldiselt lihastega ja perifeersete kudedega, leitakse hemoglobiin erütrotsüütides või punastes verelibledes (need on pseudo-rakud, st nad ei ole reaalsed rakud), mis moodustavad 40% verest.
Hemoglobiin on tetrameter, see tähendab, et see koosneb neljast polüpeptiidahelast, millest igaühel on hemirühm ja kaks kaks (kaks inimest on kaks alfa-ahelat ja kaks beeta-ahelat).
Hemoglobiini peamine funktsioon on hapniku transportimine ; teine vere funktsioon, milles osaleb hemoglobiin, on ainete transportimine kudedesse.
Tee, mis pärineb kopsudest (hapnikurikas) kudedes, kannab hemoglobiini hapnikku (samal ajal ka teised ained, mis jõuavad ka kudedesse), samas kui pöördteel on ta kudedest kogutud jäätmed, eriti ainevahetuses tekkinud süsinikdioksiid.
Inimese arengus on geene, mida ekspresseeritakse ainult teatud aja jooksul; sel põhjusel on meil erinevad hemoglobiinid: täiskasvanud inimese loote, embrüo.
Nende erinevate hemoglobiinide moodustavatel ahelatel on erinevad struktuurid, kuid sarnasusega on nende poolt teostatav funktsioon enam-vähem sama.
Mitmete erinevate ahelate olemasolu selgitus on järgmine: organismide evolutsiooniprotsessi ajal on hemoglobiin arenenud ka spetsialiseerunud hapniku transportimisele piirkondadest, mis on rikkalike piirkondadega rikas. Arenguketi alguses kandis hemoglobiin väikestes organismides hapnikku; evolutsiooni käigus on organismid jõudnud suurematesse mõõtmetesse, mistõttu hemoglobiin on muutunud, et oleks võimalik hapnikku transportida kaugemale piirkondadest, kus ta oli rikas; selleks on kodeeritud uued ahelad, mis moodustavad hemoglobiini, evolutsiooniprotsessi käigus.
Müoglobiin seob hapniku isegi tagasihoidlikul rõhul; perifeersetes kudedes on rõhk (PO2) umbes 30 mmHg: sellel rõhul ei vabasta müoglobiin hapnikku, seega oleks see hapniku kandjana ebaefektiivne. Teisest küljest on hemoglobiinil elastsem käitumine: see seob hapniku kõrgel rõhul ja vabastab selle rõhu langemisel.
Kui valk on funktsionaalselt aktiivne, võib see veidi muuta selle kuju; näiteks on hapnikuga ühendatud müoglobiin erinevalt hapnikuta müoglobiinist ja see mutatsioon ei mõjuta naabreid.
Seonduvate valkude, näiteks hemoglobiini puhul on diskursus teistsugune: kui ahel on hapnikuga seotud, indutseeritakse selle kuju muutma, kuid see modifikatsioon on kolmemõõtmeline, seega mõjutavad ka teised tetrametri ahelad. Asjaolu, et ahelad on üksteisega seotud, arvab, et ühe muutmine mõjutab teisi naaberriike isegi siis, kui see on erinevas ulatuses; kui ahel on hapnikuga ühendatud, võtavad teised tetrametri ahelad hapniku suhtes "vähem vaenulikku suhtumist": raskus, millega ahela hapnikuaatomid vähenevad, on selle kõrval hapnikuga. Sama argument kehtib ka deoksüdatsiooni puhul.
Deoksühemoglobiini kvaternaarset struktuuri nimetatakse T (pingeline) vormiks, samas kui oksühemoglobiini nimetatakse R- vormiks (vabanenud); venitatud olekus on hulk üsna tugevaid elektrostaatilisi vastasmõjusid happe aminohapete ja aluseliste aminohapete vahel, mis viivad deoksühemoglobiini jäigale struktuurile (sellepärast on "pingeline vorm"), samas kui hapnik on seotud, siis nende üksus. suhted vähenevad (sellepärast "vabanenud vorm"). Peale selle stabiliseeritakse histidiinitasu hapniku puudumisel asparagiinhappe vastupidise laenguga, samal ajal kui hapniku juuresolekul esineb kalduvus proteiini kaotada; kõik see tähendab, et hapnikuga ühendatud hemoglobiin on tugevam hape kui deoksüdeeritud hemoglobia: bohr-efekt .
Sõltuvalt pH-st seondub hemorühm enam-vähem kergesti hapnikuga: happelises keskkonnas vabastab hemoglobiin hapniku kergemini (pingeline vorm on stabiilne), samas kui põhikeskkonnas on side hapnikuga hapnikuga. tugevam.

Iga hemoglobiin vabastab 0, 7 prootonit hapniku sissetuleva mooli kohta (O2).
Bohr-efekt võimaldab hemoglobiinil parandada hapniku võimet.
Hemoglobiin, mis muudab kopsu kopsudesse kudedesse, peab tasakaalustuma rõhu, pH ja temperatuuri alusel.
Me näeme temperatuuri mõju .
Pulmonaalse alveoli temperatuur on umbes 1-1, 5 ° C madalam kui välistemperatuur, samas kui lihastes on temperatuur umbes 36, 5-37 ° C; kui temperatuur tõuseb, väheneb küllastustegur (sama rõhuga): see juhtub, sest kineetiline energia suureneb ja dissotsiatsioon eelistatakse.

On ka teisi tegureid, mis võivad mõjutada hemoglobiini võimet siduda hapnikuga, millest üks on 2, 3-bisfosfütseriidi kontsentratsioon .
2, 3-bisfosoglütseraat on metaboolne aine, mis esineb erütrotsüütides kontsentratsioonis 4-5 mM (ükski teine kehaosa ei esine sellises kõrges kontsentratsioonis).
Füsioloogilises pH juures deprotoneeritakse 2, 3-bisfosfoglütseraati ja sellel on viis negatiivset laengut; see läheb kiilu kahe hemoglobiini beeta-ahela vahel, kuna neil ahelatel on kõrge positiivsete laengute kontsentratsioon. Beeta-ahelate ja bisfosfoglütseraadi ahelate vahelised elektrostaatilised koostoimed annavad süsteemile teatud jäikuse: saadakse pingeline struktuur, millel on vähe afiinsust hapniku suhtes; oksüdeerimise ajal väljutatakse 2, 3-bisfosfütseriid.
Erütrotsüütides on spetsiaalne aparaat, mis muundab 1, 3-bisfosfütseriidi (mis tekib metabolismi abil) 2, 3-bisfosfütseraadiks nii, et see jõuab kontsentratsioonini 4-5 mM, mistõttu hemoglobiin on võimeline vahetama l hapnikku kudedes.
Kudesse saabuv hemoglobiin on vabanenud olekus (seotud hapnikuga), kuid koe läheduses on see karboksüülitud ja läbib venitatud olekusse: selles olekus olev valk on madalam kalduvus hapnikuga seonduda, võrreldes vabanenud olekuga vabastab hemoglobiin hapniku kudedesse; lisaks on vee ja süsinikdioksiidi vahelise reaktsiooni tõttu H + ioonide tootmine, seega bohr-efekti tõttu täiendav hapnik.
Süsinikdioksiid hajub plasmamembraani läbivaks erütrotsüütiks; arvestades, et erütrotsüüdid moodustavad umbes 40% verest, peaksime eeldama, et ainult 40% kudedest difundeeruvast süsinikdioksiidist siseneb nendesse, tegelikkuses siseneb 90% süsinikdioksiidist punastesse rakkudesse, sest need sisaldavad ensüümi, mis konverteerib süsinikdioksiid süsinikhappes, sellest järeldub, et statsionaarne süsinikdioksiidi kontsentratsioon erütrotsüütides on madal ja seetõttu on sisenemise kiirus kõrge.

Teine nähtus, mis tekib, kui erütrotsüüt jõuab koe, on järgmine: gradienti korral väljub HCO3- (süsinikdioksiidi derivaat) erütrotsüütist ja negatiivse laengu väljumise tasakaalustamiseks on meil kloriidide sisenemine, mis määrab osmootse rõhu suurenemise: selle varieeruvuse tasakaalustamiseks on ka vesi, mis põhjustab erütrotsüütide paistetust (HAMBURGER mõju). Vastupidine nähtus esineb siis, kui erütrotsüüt jõuab kopsu-alveoolidesse: tekib erütrotsüütide deflatsioon (HALDANE-efekt). Seega on venoossed erütrotsüüdid (otse kopsudesse) arteriaalsed.


